(по материалам доклада на 31-й Генеральной конференции ЮНЕСКО. Париж. 2001 г.).
Камакин Владислав Владимирович, Д.т.н., Маклаков Владимир Васильевич, Д.т.н.
Институт проблем управления РАН им. Трапезникова. Москва, Россия, 08.11.2021г
В данной работе рассмотрены некоторые аспекты проблемы управления состоянием живых систем (далее – ЖС) с использованием т.н. выращенных персональных белков (ВПБ) – специфического продукта, приготовленного из биомассы, получаемой культивированием клеток, взятых биопсией у ЖС, как объекта управления [1,2]. Оценка эффективности применения такого рода продукта возможна при изучении моделей ЖС, построенных на ряде следующих, феноменологически обоснованных предположениях:
1) жизнедеятельность любой ЖС обусловлена процессами метаболизма, заданными множеством соответствующих метаболических функций {U}, включающим в себя анаболические {U} и катаболические {U*} функции отвечающие за процессы, соответственно, синтеза и распада вещества ЖС.
2) ЖС так или иначе внешне структурирована; распределение важных, «опорных», структурных элементов в определенной степени отражает состояние ЖС может быть оценено методами морфометрии. Для обобщения результатов промеров, используется т.н. симилярная (от англ. similar – похожий) функция Ψ( L ), описывающая размещение измеряемых (опорных) элементов структуры в наблюдаемых областях ЖС. Данная функция имеет вид:
Ψ(x,t) = cosα [πln(1-x/L):lnΦ(t)], где: (1)
α =2L/σ; x – координата участка ЖС; L — характерный размер ЖС;
σ — область локализации элемента структуры;
На Рис.1. представлен график функции Ψ( x, t ) симилярного распределения.
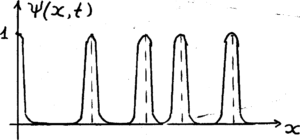
Рис.1. График Ψ(x, t) симилярного распределения
В местах фактического расположения опорных элементов структуры ЖС, функция Ψ(x,t) тождественно равна 1.
Величина Φ(t) в формуле (1) — структурный параметр, равный отношению величин расстояний между тремя ближайшими элементами структуры ЖС на момент t проведения замеров.
3) все ЖС являются в высшей степени сложными сильно неравновесными системами, с множеством обратных связей для адаптации к внешним и внутренним воздействиям и сохранения постоянства жизненно важных свойств [ 3 ]. В 1932 г. американский физиолог У. Кэннон ввел термин «гомеостаз» для определения механизмов, поддерживающих неизменное во времени состояние внутренней среды ЖС, как основу их существования. Для обеспечения стационарного состояния гомеостаза в организме человека, например, существуют четыре регулирующие системы: химическая неспецифическая (лимфа, кровь), эндокринная, нейровегетативная и анимальная нервная система. Эти системы поддерживают стационарное состояние ЖС в широком интервале внешних воздействий. Однако есть основания предположить, что в экстренных случаях могут включаться дополнительные информационно-регулирующие каналы.
Так подмечено, что животные, заболевая, прибегают к ограничению потребления пищи, как универсальному средству излечения. В процессе голодания происходит усвоение организмом собственных тканей так, что масса каждого из них, практически неизменная до режима голодания, начинает изменяться. По имеющимся данным, ограничение организма в пище до полного голодания приводит к дифференцированному изменению весовых характеристик различных органов. Патофизиолог В.В. Пашутин [4], который многие годы посвятил изучению физиологических механизмов голода, подтвердил важную закономерность (т.н. принцип приоритета): “сильные” органы во время голода существуют за счет “слабых”. Наименьшие потери несет ткань нервных центров и сердце. Ниже приведены данные о потери веса органами и тканями собаки при голодании до летального исхода (в % к исходному весу ):
Органы ткани потери в % к
исходному весу
Жир 97
Селезенка 60
Печень 53,7
Мышцы 30
Кровь 26
Почки 25,9
Кожа 20,6
Кишечник 18
Легкие 17,7
Поджелудочная железа 17
Костная масса 13,9
Нервная система 3,9
Сердце 3
Аналогичная картина дифференцированного изменения массы органов и тканей наблюдается при длительном голодании у человека [5]:
Органы и ткани Уменьшение массы (в %%)
Жировая ткань 97
Селезенка 63
Печень 56
Мускулы 30
Кровь 17
Нервная система практически 0
Процесс гомеостаза, поддерживающий, практически неизменными во времени t массовые характеристики органов ЖС — это способ реализации своего рода трансляционной t — симметрии отмеченных характеристик. В режиме голодания происходит дифференцированное изменение данных характеристик, что нарушает исходную t – симметрию своим и, как нарушение любой симметрии, «модулирующим» действием создает предпосылки для формирования информационного канала, поскольку именно нарушение исходной симметрии среды канала связи дает возможность создания сигнала в данном канале. Аналогичным образом нарушается трансляционная симметрия несущей волны в радиотехнике при модулирующем воздействии передатчика при формировании радиосигнала.
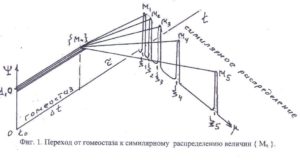
С информационной точки зрения не столь важны конкретные механизмы изменения массы Mi для i-го органа в процессе голодания. При ограничении массообмена ЖС с окружающей средой, для формирования управляющего сигнала, существенной является «симилярная» специфика изменения массы компонентов {Mn} живой системы при нарушении трансляционной t-симметрии гомеостаза (Фиг.1.):
О сигнальных свойствах белков известно ранее (Г. Блобель, Нобелевская премия, 1999г.). Соответствующие сигнальные участки и их расположение сохраняются при трансформации белковых молекул. На этом основании сигнальное свойство можноотобразить построением соответствующих симилярных функций Ψ(L) , представляющих вероятность обнаружения сигнальной метки в точке xi вдоль по длине L белковой молекулы [6].
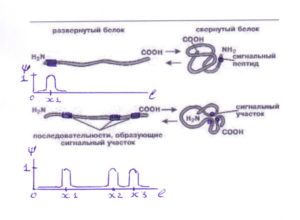
Рис.2. График симилярной функции ψ (L) распределения сигнальных меток по длине белковой молекулы ( по Г. Блобелю ).
Основываясь на изложенном, нарушение t-симметрии гомеостаза от аутолиза при голодании можно рассматривать как способ создания специфического информационного канала, по которому организм при болезни или в иной ситуации, способен оказать управляющее воздействие на составляющие его органы. Теория связи в корректной формулировке, (отмечено Винером), должна иметь дело с ансамблями сигнальных функций [7]. Как при нарушении гомеостаза, так и при учете расположения сигнальных меток Г. Блобеля эту роль могут выполнять симилярные функции {Ψ} вида:
Ψσ (Φ, m, μ) = Cosσ { π / ln [1 – U(q) ] ln [1 – μ( 1 – Φm ) / ( 1 – Φ ) ]} ( 2 )
Здесь: m – число учтенных компонентов ЖС; μ – время действия сигнального фактора ( например, аутолиза); Φ – структурный параметр, {U} — метаболические функции, ответственные за процессы анаболизма (U) и катаболизма (U*), зависящие от обобщенных переменных {q}, характеризующих состояние ЖС.
Экспоненциальные процессы синтеза и распада в ЖС представим в виде:
U ≡ U( q ) = 1 – exp (- q ln Φ ); U* ≡ U*( t ) = 2 – exp ( q ln Φ ), ( 3 )
Функция Ψ, определенная на множестве {U}, симметрична относительно преобразования U → U*, т.е.:
( U ) = Ψ ( U* ) = Ψ ( U + R ), ( 4 )
где R = ( Φ2 + Φ – 1 ) / Φ. ( 5 )
При R Ë {U} одним из условий существования ЖС будет минимизация влияния процессов и факторов R, несовместимых с метаболизмом:
R = ( Φ2 + Φ — 1 ) / Φ → 0 ( 6 )
Применение в вопросах устойчивости к ЖС вариационных методов общеизвестно [8,9]. Полагаем, что фактор R, выраженный через измеряемую величину структурного параметра Φ, характеризует степень адаптивности ЖС к воздействиям среды, а условие ( 6 ), так же как широко известные экстремальные принципы (напр.минимум диссипативной функции) определяет направление и стратегию развития ЖС. Примечательно, что минимуму значения R соответствует преобладанию в структуре ЖС значения Φ ≈ 0, 618 и преимущественному размещению опорных элементов структуры ЖС в соответствии с т.н. «золотым сечением».
Возможность использование функций (2) в качестве ансамбля сигнальных функций для формирования управляющего сигнала, при ограничении массообмена ЖС с окружающей средой, предполагает наряду с четырьмя известными системами регулирования в ЖС, появление в критической ситуации дополнительного канала регулирования, информационно обусловленного механизмом дифференцированного аутолиза собственных органов и тканей. Использование данного канала регулирования не только в режиме голодания, но и с применением ВПБ и препаратов на их основе, может быть полезна, например, для адресной доставки к органам традиционных лекарственных средств, а так же в ситуациях, о которых мы не можем сейчас судить ввиду относительной новизны рассматриваемых вопросов.
Литература:
- Камакин В.В., Маклаков В.В. Способ индивидуального подбора оптимального питания человека.Патент № 2162297 от 27.01.2001 г.
- Камакин В.В. Способ коррекции функционального состояния органов пациента. Патент № 2225211 от 17. 07. 2002 г.
- Прангишвили И.В. Системный подход и общесистемные закономерности «Синтег», М. 2000г.
- Пашутин В.В. Избранные произведения / Ред. и вступит. статья А.Д. Адо и А. М. Хомякова. — М.: Изд-во АМН СССР, 1952. — 348 с. — (Акад. мед. наук СССР. Выдающиеся деятели отечеств. медицины).
- Малахов Г.П. Голодание. Москва – Санкт-Петербург, «ДИЛЯ», 2002 г. стр. 22, 68.
- Камакин В.В., Маклаков В.В. Симилярные функции в задачах управления состоянием биологических объектов. Философские исследования. М. 2002 г. стр. 176.
- К. Шеннон Математическая теория связи.
- Левич А.П. Принцип максимума энтропии и теоремы вариационного моделирования.
- Гленсдорф П., Пригожин И. Термодинамическая теория структуры, устойчивости и флуктуаций. М., 1973г.